动作电位
动作电位(英语:action potential),指的是静止膜电位状态的细胞膜受到适当刺激而产生的,短暂而有特殊波形的跨膜电位搏动。细胞产生动作电位的能力被称为兴奋性,有这种能力的细胞如神经细胞和肌细胞。动作电位是实现神经传导和肌肉收缩的生理基础。
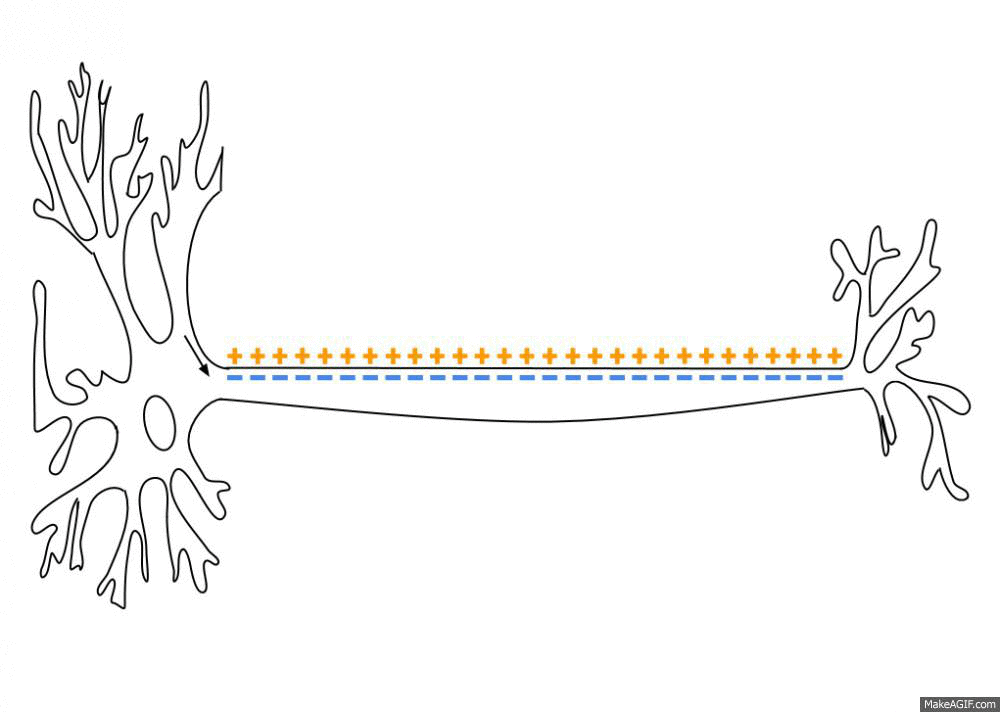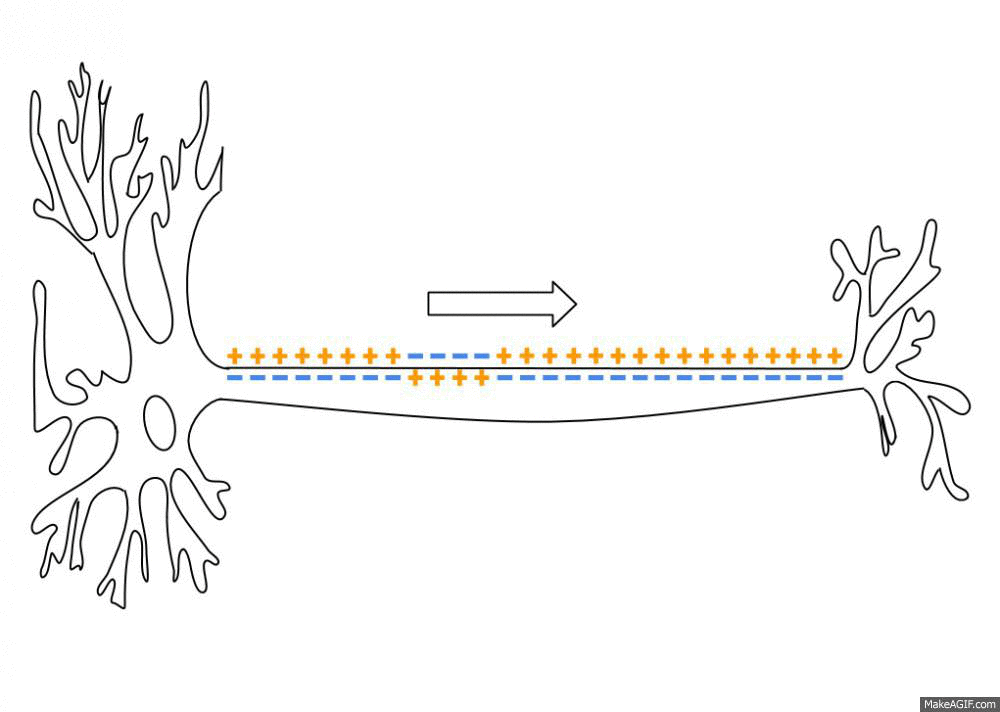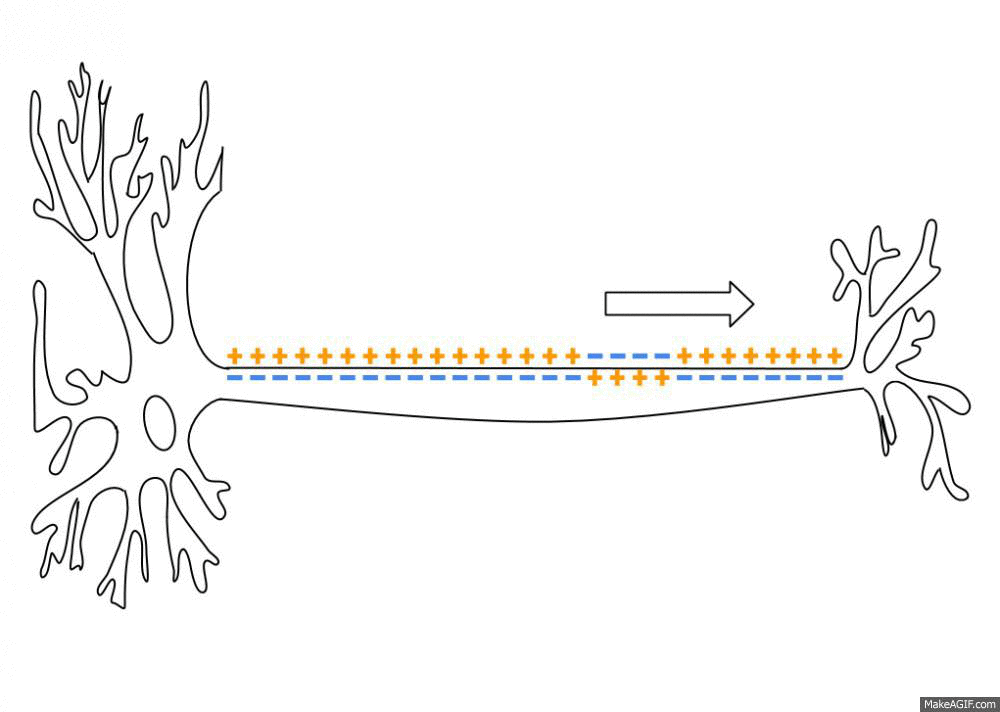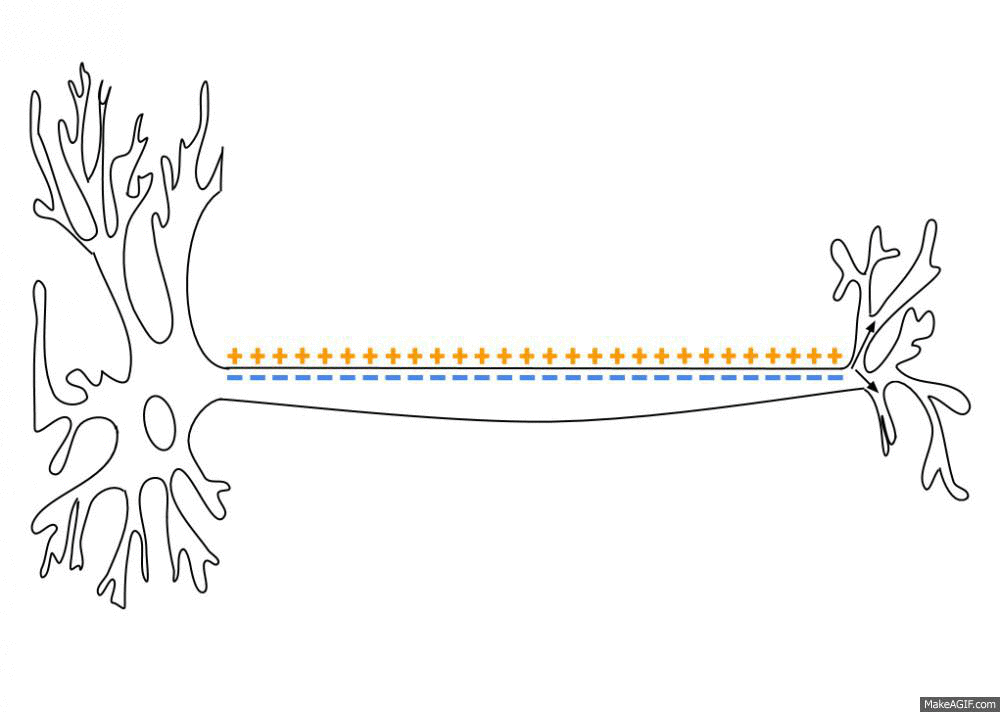
一个初始刺激,只要达到了阈电位(threshold potential),不论超过了多少,也就是全有全无律,就能引起一系列离子通道的开放和关闭,而形成离子的流动,改变跨膜电位。而这个跨膜电位的改变尤能引起临近位置上细胞膜电位的改变,这就使得兴奋能沿着一定的路径传导下去。
动作电位过程
首先细胞膜处于静止膜电位,大概在-50到-70mV。动作电位可分为4个相位:
- 极化(polarization):细胞膜由静止膜电位到达动作电位,刺激可以使细胞膜电位改变,开启电闸型钠离子通道,使钠离子大量进入细胞。
- 去极化(depolarization):膜电位陡峭上升至正值,钠离子大量进入细胞。这个“峰电位”中去极化部分被称为“升支”,而正的电位值则被称为“超射”。
- 再极化(repolarization):朝静息电位方向的下降过程。
- 过极化(hyperpolarization):再极化在下降过程中,电位会短时间下降到低于静息电位水平,然后再上升达到静息电位,这种静息电位的增大(绝对值)被称为超极化(而下降部分被称为负后电位,上升部分则是正后电位)。
动作电位持续约1-2 ms(神经元),但也可达几百毫秒(心脏)。动作电位后是不应期,这又分为0.5 ms的绝对不应期和3.5 ms的相对不应期。前者无论刺激多频繁多强都不能引起动作电位,而后者则要更强的刺激(阈电位提高了)才能引起动作电位。
动作电位的成因
关于静止膜电位的形成,请看静止膜电位。
离子通道的特征
细胞膜上有多种离子通道。而动作电位的产生,则与钠和钾离子通道有关。这些离子通道的开关状态与膜电位有关,即是所谓的电压门控离子通道。
例如钠离子通道,在静止膜电位时它是关闭并且是可激活的。当去极化到一特定值时就会引起其构象的改变,成为“打开”状态。但是离子通道却不会持续停留在开放状态,它会在几毫秒内关闭。这是通过膜上一蛋白质的失活域的活动实现的,这个失活域会像塞子一样堵住离子通道。离子通道这种状态被称为“关闭并失活的”。过渡状态关闭但可激活的只有在完全再极化后才可能出现,而开放可激活的状态是在简单模型中不可能实现的。
文献中也写道,一个关闭并失活的通道在再极化过程中首先短时间内还是开放状态,然后才改变构象直接成为关闭但可激活的。再次激活只能发生在完全再极化之后,在去极化的细胞膜中不可能存在着过渡状态开放并失活的。
当然,并不是所有的通道在电位到达一定值之时全部打开。更可能的是,通道的处于某种状态的概率是与电压相关的。而当阈电位出现时,大部分的通道便会开放,上述的模型便能很好的描述这种状态。
而状态之间过渡所需的时间也是因通道而异的。钠通道从关到开发生在1毫秒之内,而钾通道则要10毫秒。
除了电压外,还有其他开关通道的机制。对动作电位来说,有两种值得一提。一种是与“内向整流性钾通道”(Kir)有关,这种通道是不可调控的。但却有一些带正电的小分子如精素,能够在去极化到一定程度时堵塞通道孔。另一种机制与钾通道有关,当细胞间的钙离子与它结合后会开放。
动作电位的过程
{kind=link}
初始状态
初始时,细胞膜处于静止膜电位,大概在-50到-70 mV。由常开型钠离子通道、常开型钾离子通道,钠钾泵
钠钾离子决定了静止膜电位的大小进而控制离子运动的方向和强度为电化学梯度所决定。例如钠离子就有向细胞内扩散的趋向,因为其膜外浓度远较膜内高。
起始相
起始相中,要有一个刺激使得膜电位去极化到达阈值。刺激可由于突触后离子通道的开放(Na+, Ca2+)或是旁边区域产生的动作电位而形成。
当电位到达-60 mV时,Kir-通道会被精素阻断,继而导致快速的去极化并达到钠通道开放的阈值。但是外流的钾离子却会稍微阻挡这一进程(例如钙离子依赖的钾离子通道,由于在去极化过程中钙离子内流而被激活,因而会稍微阻挡这一进程)。
升支和超射
但钠通道起始要到-60 mV以上电位时才会大量开放。钠离子悬殊的浓度差,使得钠离子很快进入细胞内,造成细胞膜的去极化,更多的钠通道被激活,就有更多钠离子进入,形成了升支和正位的超射。
再极化开始
但在电位到达最大正值之前,NaV-通道已经开始失活。然后电压门控钾通道开始被激活KV。两者虽然阈值相近,但后者开放所需的时间更长,直到去极化达到一定程度才开始显示其作用。当钠内流达到最大值时,钾通道才有一半开放,而当所有钠通道失活时,钾通道的开放才到了最大值。所以钠通道开放最多的时候,是电压最大值之时,而钾通道的则位于再极化开始后。
再极化
当再极化快至静止膜电位时,KV关闭,Kir(对静止膜电位很重要)的堵塞会被去除,NaV会慢慢激活。
过极化
在很多细胞中(主要是神经元)还可以看到“过极化”。这是因为钾离子的电导性仍然高于静止膜电位,钾离子持续流出,而钠离子已停止流入,这时电位更接近钾的平衡电位(见能斯特方程)。在动作电位时,流入的钙离子会打开钾通道,这是导致钾电导性的增高的原因。当钙水平下降时,电位才会慢慢恢复正常。工作效率增高的钠钾泵对过极化也有影响。
不反应期
动作电位后,轴突在一段时间内不能再响应外界刺激。不应期的长短是由NaV活性恢复所需的时间所决定的。不应期分为“绝对不应期”和“相对不应期”。
绝对不应期紧接在动作电位后,当时再极化仍在继续,通道不能再打开。可以说,此时阈值无限大。
在相对不应期,提高刺激强度可以获得一个弱的动作电位。阈值由无限大向正常值靠近。
阈值
动作电位需要电位到达阈值才能发起,此时钠通道开放,钠离子内流。科学家尝试找出这一个阈值,但都以失败告终。神经元可以在一个范围的刺激内被激活。因此神经科学家放弃了寻找一特定阈值的努力。从系统理论来说,动作电位的产生是通过被动电位动力学和动作电位动力学的分叉实现的。但在很多教科书中,当讲解到阈值时,只会将它描述为“静息电位和动作电位间的灰色地带”。
特殊的动作电位
- 除了钠电压门控通道外,电压门控钙通道也可以引起动作电位(如心肌和浦肯野细胞)。植物细胞也能产生动作电位,例如在植物运动之时。
参见
- 静息电位(英文)一个介绍动作电位的动画
- 兴奋
- 刺激
- 运动终板
- 突触
- 神经系统
- 神经元
- 心脏
外部链结
- Das Aktionspotential (页面存档备份,存于互联网档案馆) Informationsvermittlung durch elektrische Erregung
- Alles über Aktionspotenziale
- Bildabfolge - Entstehung eines Aktionspotenzials (页面存档备份,存于互联网档案馆)
- Informationen über Aktions- und Generatorpoziale
- Elektro-Myografie (EMG) eine kleine Einführung (页面存档备份,存于互联网档案馆)
- Animation zum Aktionspotenzial